
Syracuse University Syracuse University
SURFACE SURFACE
Syracuse University Honors Program Capstone
Projects
Syracuse University Honors Program Capstone
Projects
Spring 5-1-2006
Memory & Cognition: What difference does gender make? Memory & Cognition: What difference does gender make?
Donna J. Bridge
Follow this and additional works at: https://surface.syr.edu/honors_capstone
Part of the Cognition and Perception Commons, Cognitive Psychology Commons, and the Other
Psychology Commons
Recommended Citation Recommended Citation
Bridge, Donna J., "Memory & Cognition: What difference does gender make?" (2006).
Syracuse University
Honors Program Capstone Projects
. 655.
https://surface.syr.edu/honors_capstone/655
This Honors Capstone Project is brought to you for free and open access by the Syracuse University Honors Program
Capstone Projects at SURFACE. It has been accepted for inclusion in Syracuse University Honors Program Capstone
Projects by an authorized administrator of SURFACE. For more information, please contact surface@syr.edu.
brought to you by COREView metadata, citation and similar papers at core.ac.uk
provided by Syracuse University Research Facility and Collaborative Environment
Memory & Cognition: What difference does gender
make?
Donna J. Bridge
Department of Psychology
Syracuse University
Abstract
Small but significant gender differences, typically favoring women, have pre-
viously been observed in experiments measuring human episodic memory
performance. In three studies me asuring episodic memory, we compared
performance levels for men and women. Secondary analysis from a paired-
associate learning task revealed a superior ability for women to learn single
function pairs (i.e. words that are represented in only one pair), but per-
formance levels for double function pairs (i.e. pairs that contain words that
are also used in one other pair) were similar for men and women. We also
reanalyzed data from a recognition experiment that used pictures as stim-
uli, and discovered an enhanced propensity for women to recollect the test
probes in comparison to men, but familiarity based judgments had minimal
differences between genders. A prospective study was conducted in order to
compare the effect of gender on a delayed free recall task that included basic
arithmetic problems as part of the distractor task. Implications for gender
differences are discussed with regard to biological factors involving estrogen,
and relevant social factors.

Contents
Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1
Delayed Free Recall . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2
Paired-Associate Learning . . . . . . . . . . . . . . . . . . . . . . . . . . . 10
Recognition Memory . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 19
Discussion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 25
References 32
Supported by MH069938-01 from the NIH to PI Marc W. Howard. The author acknowledges
the continuous support and assistance from Marc Howard, Jennifer Provyn, Vijay Venkatadass,
and Yaofei Zhang. Additional thanks to the remaining members of MEMlab for providing technical
and moral support.
GENDER & MEMORY 1
Introduction
Although ge nder difference s have been observed in some episodic memory do-
mains, the nature of thes e differences has not been well established or understood.
The presence of ge nder differences in episodic memory performance has been noted
under various laboratory conditions involving several different tasks, including im-
mediate and delayed free recall of words (Herlitz, Nilsson, & Backman, 1997) and
recognition of words (Herlitz et al., 1997) and faces (Herlitz & Yonker, 2002; Lewin
& Herlitz, 2002), all favoring women. The observed differences have been reliable,
but the effect sizes are often small and typically require a large subject pool to
see an effect, (Herlitz et al., 1997; Herlitz & Yonker, 2002; Lewin & Herlitz, 2002;
Kramer, Yaffe, Lengenfelder, & Delis, 2003). Nonetheless, the variable performance
accounted for by gender classification has been consistently observed in tasks me a-
suring episodic memory performance. Therefore, we find it intrinsically interesting
to the study of memory to e xamine the role that gender plays in memory processes
and its subsequent effects on performance.
By using advanced techniques to describe episodic memory processes in great
detail, we have begun to unveil subtle influences of gender. Within three episo dic
memory domains, data from four different experiments have been assessed on the
basis of gender. We compared memory performances of men and women with two
verbal learning tasks, by implementing a delayed free recall experiment and reana-
lyzing data from a paired-associate study, see (Howard, Jing, & Provyn, Submitted).
Further insight was provided by secondary analysis of two non-verbal studies mea-
suring recognition memory, that relied on the use of pictures to serve as stimuli,
(Howard, Bessette-Symons, Zhang, & Hoyer, 2006; Schwartz, Howard, Jing, & Ka-
hana, 2005).
With the present study, we attempt to describe the influence of gender that
has been observed in various episodic memory procedures, by utilizing sophisticated
methods to describe underlying processes. As a comparative feature, we examine
GENDER & MEMORY 2
the differences elicited by a basic math assessment, and provide speculation about
the potential s ource of these cognitive gender differences.
Delayed Free Recall
We are interested in mapping out the unique organization of previously learned
items or events in memory. Free recall designs provide a useful way to assess the
processes underlying episodic memory retrieval. In a free recall experiment, subjects
are presented with a list of words individually during a study phase. Following a
variable delay after the study period, subjects are instructed to recall as many words
from the list they can remember in any order. Because subjects are not required to
recall the items from the list in the same sequence they were originally presented,
the output pos itions of the reme mbered words are unique to individual subjects.
Even though there is no res triction imposed on the recall period regarding order
of output, certain patterns of remembering are consistently observed in free recall
experiments. For instance, given a list of words in an immediate free recall task, the
probabilities of recalling words located in the beginning and the end of the list are
typically greater than for words from intermediate serial positions. The heightened
probability of recall for words from early serial positions is referred to as the primacy
effect. The most often recalled words in a free recall experiment tend to be items
that occupy the final serial positions on the list. The recency effect describes this
robust property observed in free recall paradigms. Because the items located toward
the end of the list were the most recently presented, memory for these items may
be stronger than words with preceding serial positions. Generally, both of these
properties are pronounced in free recall designs; however, it is possible to reduce the
effect of primacy by prohibiting rehearsal during the task, and the recency effect
can be minimized by implementing a single delay period following presentation.
Methods. We recruited 67 women and 57 men from an introductory psychology
class at Syracuse University, all of whom participated in the study for course credit.
GENDER & MEMORY 3
We collected gender and ethnicity information from all participants, as part of the
documentation we are required to issue by the NIH. A total of 119 subjects were
used in our final analysis due to insufficient performance on the distractor task by
five subjects, discussed in further detail below. In the course of an hour, subjects
completed 18 trials of the task, each of which began with a study phase, immediately
followed with a distractor task, and finally ended with a free recall period.
For each trial, participants were instructed to study a list of 25 words, derived
from the Toronto word pool (Friendly, Franklin, Hoffman, & Rubin, 1982), and then
later asked to recall any item s from the list they could remember following a brief
delay of 30s. Each word was visually displayed on the screen for 1100 ms, in which
time subjects were required to identify the noun on the screen as a ’concrete’ or
’abstract’ word by pressing either the left or right control keys on the keyboard.
This served as an orienting task to assure the participants’ attention to the task.
The inter-presentation interval (IPI), the lapse of time in between the presentation
of each word, was set to 200ms. During the delay, subjects were prompted with
a series of basic arithmetic problems involving the addition of three single positive
integers. Answers were typed in using the number keys on the keyboard, and the
number of math problems varied with each trial depending on the subject’s response
time for each equation. After the distractor period, participants were prompted with
a beep that indicated to them that they should begin recalling any items from the
list by speaking the words into the headset provided. Each recall period lasted 60s.
Completion of the 18 trials was followed by a surprise final free recall period, during
which time participants attempted to recall any words from any of the lists that had
been presented to them during the experiment. Five minutes were allocated to the
final free recall period.
Results. Several measurements are typically used to assess patterns of learning
in free recall experiments. The first measurement we used to assess pe rformance was
a serial position c urve (see Figure 1). A serial position curve clearly demonstrates
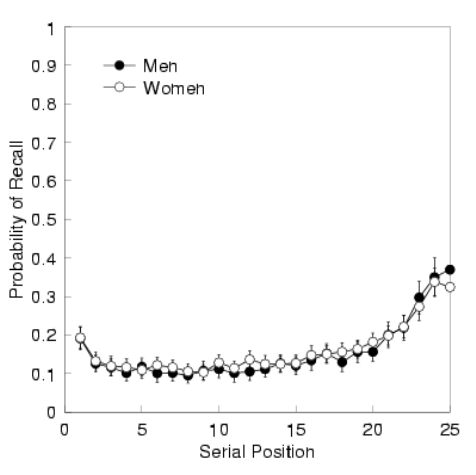
GENDER & MEMORY 4
Figure 1. Serial position curve for men and women. Error bars reflect 95% confidence
intervals.
the impact of recency and primacy on the recall period, by depicting the probability
of recalling an item from the list relative to its serial position. Not only does the
serial position curve highlight the recency and primacy effects, but it also serves as
an overview to examine the probability of recalling items distributed across all se-
rial positions. Figure 1 displays the cumulative probability of correctly recalling an
item from a list collapsed across trials. Using the serial position curve, we are able
to determine the average number of recalls per trial, by adding up each subjects’
probability of recall for every se rial position. We observed minimal differences be-
tween average number of recalls, with men recalling approximately 3.9 and women
averaging 4.03 words per trial, t(117) = −.62 p > .5.
Although the serial position curve is a useful tool to analyze free recall data,
it is limited in its ability to unveil subtle patterns of retrieval. A graph of the prob-
ability of first recall is useful for providing additional insight into the complexities
that contribute to free recall paradigms. The probability of first recall (PFR) graph

GENDER & MEMORY 5
Figure 2 . Probability of First Recall. Error bars indicate 95% confidence intervals.
is essentially a serial position curve, but it only considers the very first item re-
called, and neglects subsequent output positions (Howard & Kahana, 1999, 2002).
In immediate free recall, the PFR graph reinforces the pronounced recency effect
depicted in standard serial position curves. Its main contribution, however, is its
novel ability to unveil the heightened probability of recalling items from the end of
the list in early output positions. The PFR curve (Figure 2) depicts similar results
to the serial position curve, with men and women demonstrating indistinguishable
probabilities of recall for the first output position.
It is possible to attenuate the pronounced recency effect observed in immediate
free recall by increasing the lapse of time between presentation and recall, while
simultaneously implementing a distractor task to disrupt rehearsal. The diminished
recency effect can be observed in either the PFR or the serial position curve. Both
curves highlight the primacy effect that remains in a free recall task with a delay, and
it displays a reduced recency effect that is comparable in size to the primacy effect.
We used a delayed free recall task in order to divert subjects’ tendency to mainly

GENDER & MEMORY 6
Figure 3. Conditional Response Probability curve for men and women. Error bars are 95%
confidence intervals.
recall words from the end of the list, so that we would be able to focus primarily
on the transitions that occur between items dispersed throughout the list during
recall. An additional measurement is used to analyze specific transitions between
words according to their output positions and relative to original serial positions
at which they were presented. We constructed a conditional res ponse probability
(CRP) curve to describe the temporal relationship between words from the list with
consecutive output positions. The CRP curve captures the unique organization of
items in memory based on the two properties of contiguity and as ymme try. Figure 3
shows the probability of recalling an item from the list set as a function of lag, where
lag is the relative difference between serial positions of subsequently recalled items.
So, the relationship between items that are recalled with succeeding output positions
is depicted by the distance be tween their serial positions on the list. A small lag
between successively re called items means that they were originally presented in
similar contexts of occurrence based on time and location. Items recalled with
ensuing serial pos itions are denoted by positive lags, while negative lags represent
GENDER & MEMORY 7
preceding items on the list. For instance, suppose a subject is presented with a list
of words, denoted here by letters, A - B - C - D - E - F. During the recall period, if
the subject responds with D and then follows that recall with E, the lag would be
indicated by +1. If the s ubject recalled C after D instead of E, the lag would be
assigned a −1.
The weight of the associations presumably made during the encoding state
are revealed during retrieval, and rely heavily on the amount of c ontextual overlap
between the items. This is clearly demonstrated by the heightened conditional
response probability of items with small absolute lags and a diminishing probability
of recall as the lag increases. Interestingly, the CRP curve also exposes the peculiar
property of asymmetry of associations. The CRP curve reaches its maximum value
with a lag of +1, while a lag of negative −1 yields a drastically diminished probability
of recall, but its value is still relatively high in comparison to probabilities with
much greater absolute lags. The CRP curve (see Figure 3) provides the probability
of recall relative output position, collapsed across output positions. We compared
probabilities of recall with a lag of +1 and −1 but did not observe any distinguishable
differences.
We examined the data from the delayed free recall experiment using a serial
position curve, PFR curve, and CRP curve. These three tools permitted us to
describe some the intricacies that underly memory retrieval. We compared men’s
and women’s performances, but we did not observe any significant effects of gender
with any of the measurements we utilized for this study.
Math Distractor. We initially examined the distractor analysis to confirm that
subjects were following the directions of the task and not simply spending the 30s
delay rehearsing words from the previous list. Our analysis contained several mea-
surements describing each subject’s performance, including the total number of math
questions answered correctly and incorrectly, overall proportion correct, average re-
action time, and variable reaction time. We decided to exclude those participants
GENDER & MEMORY 8
who were not appropriately completing the task. Participants that exhibited vari-
able reaction times greater than three standard deviations from the overall mean
(> 4989 ms) were cut from the analysis, as well as those whose proportion correct
was more than three standard deviations away from the mean (< .77 correct). A
total of five subjects (all men) were excluded from our final analysis.
Interestingly, we discovered significant gender differences in the math distrac-
tor task. We used t-tests to compare reaction times, proportion correct, and total
number of correctly answered equations. Overall, men and women answered the
same proportion of math problems correctly (.93 vs .92respectively), t(117) = 1.18
p > .2. Even though percentage correct was nearly equivalent, we discovered large
advantages for men regarding total number correct and mean reaction time. Men
were able to answer significantly more equations correctly t(117) = 3.05 p < .003,
and at a faster rate t(117) = −2.4 p < .02, than women. Figures 4a and 4b display
the distribution of total correct responses for men and women.
Distractor Discussion. Even though the cumulative number of questions an-
swered correctly was higher for men, the level of accuracy was equivalent for both
groups. Men may have been able to answer more math equations overall because
their mean response times were significantly faster than women’s. The ability to cor-
rectly answer basic math problems does not seem to differ across genders; however,
since response time is a standard of meas urement that is considered when assessing
performance levels, men appear to be superior on this particular mathematical task.
Our examination of the detailed distractor analysis led to an unexpected in-
fluence of gender. The implementation of a distractor task in a delayed free recall
experiment is primarily to divert subjects from rehearsal. The magnitude of the
distraction produced by the task should be relatively equal among participants. Be-
cause of the divergent influence of gender on performance for the math task, we
cannot assume that both groups were equally affected by the distractor task, and
therefore men’s and women’s performances on the memory task may have been dif-

GENDER & MEMORY 9
a
b
Figure 4. Distribution of total number of correct responses made by men and women. a.
Distribution for men. b. Distribution for women.
ferentially influenced by the math problems. In future memory studies that do not
take gender into consideration, the type of distractor task should be reevaluated so
that all participants are comparably distracted.
The prevalence of stereotype threat may have an impact on some women’s
math performance. Stereotype threat refers to the psychological threat experienced
by individuals who fear being categorized by negative stereotypes of a group they
are affiliated with (Pronin, Steele, & Ross, 2004). Stereotype threat has the ability
to affect performance of individuals in such a way that it ac tually reinforces the neg-
ative stereotype associated with the individual’s group. Findings from some studies
have suggested that mathematical evaluations can induce the negative influence of
GENDER & MEMORY 10
stereotype threat on women’s performances (Pronin et al., 2004; Johns, Schmader,
& Martens, 2005). Presumably, the stress evoked by stereotype threat may provoke
women to doubt their mathematical proficiency, causing them to exhibit slower re-
sponse times than men. This stress may be a consequence of the broadly supported
assumption that men are naturally more inclined to succeed in the realms of math
and science.
The distractor analysis provides interesting insight into the standards by which
we measure performance. For instance, standardized tests presumably measure
mathematical ability and comprehension, but the strict time constraint imposed on
these examinations may contribute to the divergent test scores exhibited by men and
women. Because mathematics is an explicitly gendered domain of academia, where
the achievements of men are greatly encouraged and a high aptitude is necessarily
predicted at a very young age, women are subject to oppositional group stigmas
(Pronin et al., 2004; Johns et al., 2005). Consequently, women who are affected
by this stigmatization may demonstrate performances that reflect presupposed ex-
pectations about their individual abilities that ste m from negative generalizations
implicated by their particular gender group affiliation. The additional stress ex-
perienced by some women may induce doubt and anxiety that is exposed by their
comparatively lengthy response times in relation to men.
Paired-Associate Learning
Understanding the underlying processes that determine memory performance
is essential to break down the observable differences in memory between genders.
Rather than measuring episodic memory performance by solely focusing on men’s
and women’s aggregate correct recalls in verbal learning tasks, our analysis has been
primarily devoted to the mechanisms underlying the retrieval of concrete informa-
tion. In order to do this, we have assessed the learning of words by examining the
associations formed between items by using a paired-associate learning task.
GENDER & MEMORY 11
Associative memory can be described on multiple levels by assessing the le arn-
ing of two pair types, double function and single function pairs. Single function pairs
are compose d of unique words that are used in only one pair, as seen in Figure 5 a.
Double function pairs consist of words that are used in two separate pairs (Primoff,
1938), as seen in Figure 5b. The words used in the double function pairs are part of
a linked list; each word is matched with two unique item s from the double function
list and serves as a stimulus for one word and a response for another word during
the test phase. For example, C-D and B-C are two pairs that share a similar item,
C, that plays a specific role in each pair; as the correct response for the cue B and
the probe for item D. Two particular pairs on the list do not follow the constraints
of either pair type; the first and last pairs on the double function linked-list. The
first and last pairs on the double function list A-B and H-I are distinguished from
the other pairs, because A and I are represented in only one pair. Therefore, the
learning of these two pairs is disregarded from our analysis of single function and
double function pairs.
In this type of experiment, subjects are randomly presented with a list con-
taining both pair types. Pairs are presented in an arbitrary order, however double
function pairs containing a shared item are never presented consecutively. This
constraint provides a means to assess the construction and strength of associations
formed without the presence of direct temporal proximity between items in separate
pairs. Because each double function item is a member of two separate pairs, recall
is more difficult for this pair type than it is for single function pairs. This is pre-
sumably due to the presentation of each double function item in two distinct, yet
overlapping contexts of occurrence (Howard et al., Submitted).
Intrusions. Our analysis takes into account all possible res ponse s made to a
cue item, including correct recall and intruding items. Although there is only one
correct response to each stimulus, there are several different types of intrusions one
could make in response to a double or single function item probe. The only other

GENDER & MEMORY 12
a b
A B C D E F G H I
Figure 5. This example is representative of a list of words depicted by letters. Each item
on the single function list is a novel word used in only one pair, while maintaining a solitary
serial position relative to the pair. Double function items are presented in two separate
pairs with differing serial positions within each pair. a. Single Function List. b. Double
Function List.
possible circums tance that arises during the presentation of the cue is when the
subject do e s not elicit any response to the stimulus. In this case, we simply char-
acterize their error as no response and do not assimilate these errors with intrusive
responses. The correct response to a cue, C (se e Figure 5b), would be its corre-
sponding pair member, D. A response of B would indicate a backward intrusion.
Even though B and C do share group membership in a different pair, the order of
presentation of the words in the pair is indicative of the correct response. The dual
presentation of items in two separate pairs incites confusion, and often times lures
participants to make incorrect responses. A remote intrusion to the cue C, could
actually be one of several responses, including E or A, for example. Other than
the backward intrusion and the correct response, a remote response embodies all of
the words that are members of the same pair type as the cue, such as I or F. An
other response can be thought of as being the functionally opposite response to a
cue as the remote intrusion. Rather than being an inconsequential response to a
cue from the same pair-type list, an other intrusion is any item that is a m embe r of
the opposite type of pair, for instance J or R , in response to the cue C. The final
type of intrusion one could make is referred to as an extra-list intrusion, which is
reasonably self-explanatory, in that the intruded item does not come from either of
the lists, for example Z. The type of responses one could make to a single function
probe follow similar guidelines to those constructed for a double function cue as in
the examples above, but the unique properties of those responses will be described
in greater detail below.
GENDER & MEMORY 13
We broadly characterize all incorrect responses to a probe item as associative
and non-associative intrusions. The intrusions falling under the category of associa-
tive intrusions include backward and remote responses. One theory attributes the
formation of associations between items to overlapping temporal contexts of occur-
rence (Howard & Kahana, 2002; Howard et al., Submitted). A backward intrusion
demonstrates this tendency clearly. For instance, B (see Figure 5b) was originally
presented in two different pairs with overlapping contexts. A backward intrusion
would presumably be a likely response to a double function item, due to its strong
contextual and temporal overlap with the probe (e.g. B given C ). Remote responses
are also considered to be associative intrusions, even though these intrusions were
never actually presented with the probe item in a pair. Since a temporal context
is retrieved when prompted with C, items that share a context or reserve a strong
overlapping context, indicated by a small linked-list lag, are more likely to be re-
called than items that have a weaker contextual overlap with a large linked-list lag,
or items with no overlap at all (Howard e t al., Submitted).
Single function cues also induce remote and backward intrusions, but their
connection to the probe is not so obvious. A sequence of single function pairs is
arbitrarily constructed to form the single function list, and then the list is presented
in a randomized order. Therefore, backward intrusions are not provoked by the
sharing of common items during presentation. Instead, a backward intrusion indi-
cates a lag of −1 from the cue item on the serial position list that was constructed
for the purposes of making comparisons across lists. Because the single function
pairs are not members of a linked-list like double function pairs, a remote intrusion
does not involve any mediating items that facilitate direct c onnections across pairs.
These intrusions are defined by any incorrect response from the single function list
to a single function pair, excluding the backward response. Remote responses are
potentially provoked by their similar membership with the same pair type.
Responses with no contextual or temporal overlap are considered to be non-
GENDER & MEMORY 14
associative intrusions. These intrusions can be similarly explained for double func-
tion and single function cues. An other intrusion refers to any item that was orig-
inally presented as part of the oppositional list type (i.e. single function vs double
function) of the cue. Another type of non-associative intrusion one could make is
an extra list intrusion, which is indicative of a res ponse that was not initially pre-
sented to the subject as part of the study list. No response by a participant is
self-explanatory and implies a non-associative intrusion, but this type of response is
not included in our analysis.
Methods. We conducted secondary analyses on a large paired-associate data
set from a study reported in Howard et al. (Submitted). We divided the data by
gender in order to examine its effect on the learning of double function and single
function pairs. Gender was determined for the purpose of our analysis by referring
to documentation collected at the time of the study. Two-hundred-six subjects,
147 women, were used for the gender analysis out of a total of 216 subjects who
participated in the study. Three subjects’ data were excluded from the analysis due
to lack of paperwork indicating gender specification, and seven subjects (5 women)
were cut because they did not properly complete the task, as indicated by their
failure to meet the criterion of rece iving a probability of correct recall > 0 for all
trials of either pair type.
For a complete description of the me thods used in this task, please refer to
Howard et al. (Submitted).
Results. We measured the probability of correct recall and intrusions for both
gender groups on each of the four trials. Comparisons were made between double
function and single function pairs to examine learning across trials and frequency
and type of intrusions. See Table 1 to view proportions of correct recall and intrusion
types for single function and double function pairs across genders.
In order to consider the impact of gender on associative learning, we examined

GENDER & MEMORY 15
Single Function
Men Women
trial cr bk rm othr xli nr cr bk rm othr xli nr
1 .299 .002 .043 .219 .096 .341 .369 .003 .037 .190 .068 .333
2 .601 .002 .045 .149 .060 .143 .642 .001 .033 .128 .060 136
3 .716 .002 .018 .094 .072 .098 .748 .003 .020 .087 .050 .093
4 .761 .000 .016 .082 .062 .080 .792 .005 .022 .069 .049 .063
Double Function
Men Women
trial cr bk rm othr xli nr cr bk rm othr xli nr
1 .257 .158 .204 .043 .065 .273 .259 .180 .199 .035 .053 .273
2 .386 .181 .149 .038 .060 .185 .415 .201 .159 .031 .044 .150
3 .490 .166 .118 .027 .054 .145 .496 .188 .133 .023 .044 .116
4 .540 .152 .107 .024 .052 .125 .546 .170 .121 .023 .039 .101
Table 1: Proportion of correct recalls and intrusions. See text for a complete descrip-
tion of responses, cr: correct recall; bk: backward; rm: remote; othr: other; xli: extra-list;
nr: no response
differences in probability of correct recall across trials and pair type. Figure 6 shows
the proportion of correct recalls made by men and women for single function (Figure
6a) and double function (Figure 6b) pairs separately. It is clear from the picture that
women outperform men on single function pairs, but they appear to have similar
levels of performance when probed with a double function stimulus. We tested
for significance by using a three-factor within-subjects ANOVA, with probability
of correct recall se t as the dependent variable, and trial, pair type, and gender
set as independent variables. Consistent with previous findings (Howard et al.,
Submitted), a significant main effect of trial, F (3, 1498) = 684.388, MSE = 11.061,
p < .001, pair type, F (1, 1498) = 1049.848, M SE = 16.967, p < .001, and an
interaction of trial and pair type F (3, 1498) = 36.590, M SE = .591, p < .001
were observed. Interestingly, a new finding illustrated by the ANOVA revealed a
significant interaction of pair type and gender F (1, 1498) = 6.2982, MSE = .102,
p < .05.

GENDER & MEMORY 16
a b
Figure 6. Probability of correct recall across trials for single and double function pairs.
Error bars reflect 95% confidence intervals. a. Single Function. b. Double Function.
Associative Interference. The interaction of gender and pair type is evidence of
women participants’ superior ability to correctly learn single function pairs of words
(Figure 6a), but not double function pairs (Figure 6b). Based on womens’ superior
performance on single function pairs, we would expect the difference between mens’
and womens’ probability of correct recall to remain consistent across pair types.
However, men and women maintained an approximately equivalent probability of
recall on double function pairs across trials.
These results suggest that women are more susceptible to associative inter-
ference than men on double function pairs. Perhaps women were prohibited from
achieving a higher probability of correct recall in relation to men, because their
memories elicited other items on the double function list that were temporally and
contextually associated with the probe. For instance, if women were more likely
than men to remember a backward ass ociated item to a cue, it may prevent them
from delivering the correct response.
Given women’s superior performance on single function pairs, we predict that
GENDER & MEMORY 17
they are more susceptible to associative interference than men. If women do exhibit
more assoc iative responses than men to double function cues, this would reasonably
explain why their probability of correct recall is comparatively lower than one would
expect.
In order to investigate this possibility, we analyzed the intrusion data. We
limited our intrusion analysis to incorrect responses that came from the study list,
and ignored extra list intrusions and no responses. This strategy made it possible
to assess specific transitions made between items on the list.
Figure 7 depicts the proportional relationships among the backward, remote,
and other responses for s ingle function (Figure 7a) and double function (Figure
7b) cues, given that the subject made an intrusion from the study list. The s ingle
function intrusion data seems jumbled and unsystematic, and without the presence
of any apparent gender difference s. Additionally, the intrusion data was limited for
single function cues, because of the heightened probability of correct recall (see Ta ble
1), resulting in minute values assigned to the remaining responses. The plots from
the double function intrusion data are comparatively systematic and organized, with
distinguishable gender differences. When an error is made from the list in response
to a double function cue, women seem to be making more backward and remote
responses than men. This can be seen by the systematic displacement downward
of women’s data plots, that is seemingly drawing them away from other intrusions,
and pulling them closer to backward and remote responses.
To quantify Figure 7b, we tested for significance using a two-factor ANOVA.
We constructed an associative index for each subject that compared the probability
of making an associative intrusion versus a non-associative intrusion from the list
(i.e. the probability of making a backward or remote response minus the probability
of making an other response, divided by the combined probabilities of the three
intrusions). We utilized this technique in order to control for inconsistent probabil-
ities of recall and no response rates across genders for both pair types. Using the

GENDER & MEMORY 18
a b
Figure 7. Proportion of backward, remote, and other intrusions in 3-D space, when all other
types of responses are excluded. Bk + Rm + Othr = 1. a. Single Function. b. Double
Function.
associative index as the dependent variable, and trial and gender as the indepen-
dent factors, we observed a main effect of gender F (1, 846) = 6.3125, M SE = .252,
p < .05 for double function cues, as we had predicted. The associative index from
the single function intrusion data did not result in a significant effect of gender,
F (1, 480) = 1.9736, MSE = .796, p > 0.1.
In response to a double function probe, women were significantly more likely
than men to make an associative intrusion from the list rather than a non-associative
response from the list. Even though m en and women were seemingly performing
at the same level on double function pairs according to their equal probability of
recalls, women were able to better remember the context of the probe when they
were unable to recall the correct answer. Presumably, women were more subject to
the associative interference brought on by a double function cue, leading to a higher
proportion of backward and remote intrusions in comparison to men. Somewhat
counterintuitively, in this particular case , a better memory is reflected by excessive
errors.
GENDER & MEMORY 19
Recognition Memory
We initiated the examination of secondary analyses from two non-verbal tasks
on the basis of gender in order to gain additional insight into the specificity of the
gender differences that have previously been observed. By solely relying on the
visual display of pictures to provoke learning, we compared recognition memory for
men and women.
Recognition tasks differ from recall tasks, in that subjects are not expected
to independently produce information from memory. Rather, recognition studies
require participants to discriminate be tween items that have previously bee n studied
and novel stimuli that were never b e fore presented. A great deal of controversy exists
in the field of cognitive psychology about the influences that determine recognition
memory. Traditionally, dominant theories have attempted to describe recognition
memory as being dependent upon a single process of familiarity, where responses are
made on the basis of some threshold of confidence that subjects use to discriminate
between “old” and “new” items (Yonelinas, 2001). However, in recent years more
researchers have attributed episodic recognition memory to the contribution of two
distinct components: recollection and familiarity (Yonelinas, 2001). Recollection
can be thought of as a process synonymous to recall, where c orrect identification of
an item relies upon the specific properties associated with the item and a concrete
link to the original presentation of the item (Yonelinas, 2001; Schwartz et al., 2005).
Other judgments made during a recognition study are thought to rely on the notion
of familiarity, where responses are influenced by the similarity of the probe item and
information previously stored in memory (Schwartz et al., 2005).
The rather intuitive approach of dual-process recognition models can be
demonstrated in real-life situations. Suppose you reflect upon your commute to
work last Friday morning. You may vividly recall walking in late at 10 : 00a.m.,
because you missed the 9 : 15a.m. bus. During your stroll, you noticed the baby blue
sky, illuminating sunshine, and cool breeze on this seasonally warm day in April,
GENDER & MEMORY 20
that caused you to put on your sunglasses and carry your sweater. Your ability to
recall your morning walk with a great deal of specificity about the time and con-
text of the event would be classified as a recollected memory. On the other hand,
imagine that you were asked to recall your commute to work last Friday, but a vivid
description of your trip may not come to mind. You may m ake a few as sumptions
and generalizations about the transit to work that day. For instance, you know
where you were employed at the time, you can vaguely remember missing the bus,
but you do not recall how you actually got to work that day. Even though some
general information is available after some rumination, you are unable to determine
precisely the course of events that shaped your commute to work on that day. This
type of generalization would be characterized by its familiarity-based nature. You
are not able to restructure the event with spec ific information from that day, but you
can make a reasonable estimate about the trip to work that day based on general
knowledge and information you have maintained from that period of time.
We measure recognition performance by first classifying the types of responses
one could make to a stimulus. In a recognition task that assesses memory of pictures,
the subject must identify if (s)he had previously studied the item or if the picture
was new to the task. If the cued picture was in fact an “old” item from the list, the
subject could essentially make one of two responses. If the subject correctly identifies
the item as “old”, the response would be considered a “hit”. However, if the subject
mistakenly rejects the “old” item as “new”, the response would be categorized as
a “miss”. Similar to the presentation of an “old” item during the test phase, if
the subject is probed with a “new” item, there are two responses (s)he could issue.
A “correct rejection” yields the appropriate answer, while a “false alarm” suggests
that the participant misattributed the “new” for an “old” item. Because “hit” rate
predicts “misses”, and “false alarm” rate specifies “correct rejections”, we primarily
focus on these two types of responses when constructing descriptive figures for the
data.
GENDER & MEMORY 21
Using a dual-process approach, we attempt to measure recognition memory
performance and classify decisions as either recollection or familiarity based judg-
ments. The model is particularly useful when describing receiver operating charac-
teristics (ROCs) and making comparisons across groups. A ROC graphs the propor-
tion of correctly identified “old” items (hit rate) set as a function of the proportion of
incorrectly recognized “new” items (false alarm rate) (Yonelinas, 2001). The graph
is constructed by using a variable number of criteria set for the task. Rather than
simply measuring “sure old” and “sure new” responses for each probe, recognition
tasks provide a gradient of response criteria. This way, subjects are able to convey
their level of confidence for each stimulus presented. The farthest left point on the
ROC denotes the most stringent criteria, whereby the highest confidence responses
are made. Each subsequent plot to the right of the first point include the cumula-
tive values of less and less confident responses (Yonelinas, 2001) (see, for example,
Figure 8. So, the hit rates and false alarm rates are relationally determined. In
an experiment with six criteria, a response of 6 for an “old” item will always be a
hit, while a response of 5 to an “old” item will be the cumulative proportion of hit
rates to false alarms for responses 5 and 6. As the criterion becomes more and more
liberal, the ROC approaches the maximum values of both probabilities (1, 1).
The Yonelinas dual-process high threshold model utilizes two free parame-
ters to generate an ROC; discriminability d
0
and recollection R. Discriminability
is computed as the average increase in familiarity assigned to items. This param-
eter measures the difference between the equal familiarity distributions of “new”
and “old” items. Recollection is determined by the probability of recollecting an
“old” item from the study list (Yonelinas, 2001). We use these two parameters to
assess recollection and familiarity across genders using methods described in detail
in Howard et al. (2006).
Methods. Analysis from two similar recognition tasks were assessed in order to
determine gender differences in recollection and familiarity. This secondary analysis
GENDER & MEMORY 22
was conducted after dividing the data based on gender identification specified at the
time of the original studies.
Both studies used travel pictures as stimuli, taken from planetware.com, a
travel picture website, and included a variety of scenes from throughout the world
(Howard et al., 2006; Schwartz et al., 2005). The criteria to ass es s confidence levels
was set to 6, so participants had six choices to use in response to one item. A
response of 1 would indicate that the subject was sure the stimulus was a “new”
item that had not been studied. Responding to an item with a 6 would indicate
that the subject was sure it was an “old” item that had definitely bee n studied.
Responding with one of the numbers in between the extreme values of 1 and 6
reflect less confident responses (Howard et al., 2006; Schwartz et al., 2005).
Experiment 1. The analysis from the first experiment included data from 19
women and 23 men. A detailed des cription of the experimental methods can be
found in Howard et al. (2006).
Experiment 2. The secondary analysis from experiment 2 used data from 49
women and 21 men. A complete description of the experimental methods from this
study can be seen in Schwartz et al. (2005).
Results. We used a dual process model to measure recognition mem ory perfor-
mance and describe ROC curves. The estimated roles of recollection and familiarity
can be observed by the curvature and shape of the ROCs. In the YHT (Yoneli-
nas, 2001), if familiarity were the only process contributing to responses, ROCs are
constructed that are symmetric along the diagonal line. If responses reflect some
recollection based decisions, the leftmost point on the ROC will be s hifted up, result-
ing in asymmetrical lines along the diagonal. Since the contribution of familiarity
and rec ollection vary, the shape of the ROCs do not completely depend on levels of
accuracy (Yonelinas, 2001). Although other approaches to fitting ROCs are possi-

GENDER & MEMORY 23
a b
Figure 8. Receiver Operating Characteristic (ROC) curve, with hit rate (HR) set as a
function of False Alarms (FA) shown for two recognition experiments. a. Exp eriment 1. b.
Experiment 2.
ble, here the YHT provides superior fit to ROC curves from pictures (Howard et al.,
2006).
The differences in familiarity and recollection contributing to men’s and
women’s memory performance can be assessed by examining ROCs generated from
experiments 1 and 2, see Figures 8a & 8b. Interestingly, the two independent com-
ponents of recognition memory were not equally influenced by gender. The ROCs
for men and women are similarly curved, but they elicit noticeable differences. Even
though the shapes are relatively similar, the women’s curves are clearly raised above
the men’s for high confident responses in both experiments. As the criterion be-
comes increasingly liberal, the distance between the lines is diminished. The graphs
from the two recognition studies suggest that females have a higher probability of
R, but levels of familiarity are approximately the same for men and women.
By applying the parameters provided by the model to our data, we were able to
quantify the subtle differences observed from the ROCs. We compared recollection
R and discriminability d
0
results from each task across gender groups by using t-tests.

GENDER & MEMORY 24
Experiment 1 & 2 Mo del Parameters
R d
0
Exp Men Women Men Women
1 .27(.03) .35(.04) .55(.08) .55(.07)
2 .36(.03) .41(.02) .44(.06) .55(.05)
Table 2: Model Parameters for recognition experiments 1 & 2. The standard
error of each mean is in parentheses.
Assessment of the individual experiments separately did not yield a main effect of
gender. This may be a consequence of the small subject pools allocated for each
experiment, combined with the consistently small, but robust gender differences
that have previously been observed in episodic memory tasks. In order to minimize
the variability provoked by a small number of subjects in each gender group, we
combined the results from experiments 1 and 2. The collective analysis was assessed
by use of a two factor ANOVA with experiment and gender set as independent
variables. Significant main effects of experiment F (1, 109) = 6.603, MSE = .155,
p < .05 and gender F (1, 109) = 5.469, M SE = .128, p < .05 were observed when we
considered R as the dependent variable. We did not observe significant differences
in discriminability for gender or experiment.
Because the assessment of recognition memory is not necessarily straightfor-
ward, we have relied upon a model to describe performance across genders. We
relied upon the parameters of the YHT dual-process model (Yonelinas, 2001) to
quantify the gender difference s from the recognition studies. We have used this
model as a mechanism to substantively describe the gender differences we observed
from the actual tasks. These findings from these two studies suggest that men and
women rely equally on familiarity to make judgments. However, measurements of
recollection, presumably a function that is dependent upon a specific memory of an
event, are indicative of reliable gender differences favoring women.
GENDER & MEMORY 25
Discussion
The intricacy of our reliable measurements have allowed us to the examine the
impact of gender on episodic memory processes in great detail. Memory performance
from the delayed free recall design did not differ across genders; however, a large
gender effect was indicated by the math distractor task. Although discreet gender
effects have been observed in some declarative memory tasks, we have neglected
to identify an influence of gender that is of the same magnitude as the difference
yielded by performance on the mathematical assessment.
The paired-associate task provided interesting findings regarding gender. Al-
though women were able to correctly respond to a single function cue more often
than men, double function probes did not elicit significant gender differences in
probability of correct recall. Presumably, women did not surpass the performances
of men on the double function cues because they were more susceptible to associative
interference, indicated by their higher probability of responding with an associative
intrusion to a double function probe in relation to men.
The two recognition studies, which assessed memory performance with the use
of visually displayed travel pictures, elicited similar findings. The ROCs from the
individual experiments clearly depict women’s superior ability to recollect pictures,
but familiarity for both genders is relatively indistinguishable.
Studies involving gender effects on cognition have typically relied upon two
schools of thought to explain the source of the observed differences. The expla-
nations occupy either the biological realm or they tend to favor social factors as
being the primary source of influence. It seems likely that both social and biological
factors play a role in generating differences between sex categories. Because direct
connections between neurological m odeling of the brain based on animal studies
and cognitive models involving human behavior cannot be made, speculative im-
plications can only be considered. Established biological sex differences cannot be
simply translated into gendered cognitive differe nces , because these characteristics
GENDER & MEMORY 26
are influenced by social and biological factors.
It is first essential to bear in mind several confounding factors that may have
influenced the results from our secondary analysis. Unlike the delayed free recall
task we designed and conducted, the data from the recognition and paired-associate
studies was examined on the basis of gender after it was originally collected. There-
fore, we were unable to control for several variables that typically arise during the
course of the semester. For instance, the quality of our subjects varies throughout
the semester, because the majority of our subject pool consists of students who are
taking an introductory psychology class and volunteer to participate in our research
studies for course credit. Those subjects who illustrate the tendency to procrasti-
nate in fulfilling their research credit and participate toward the end of the semester
usually do not provide the most useful data and may not match the caliber of the
subjects who participated earlier in the sem este r. Although this is not necessarily
the case, it could be confounded with gender.
Another factor to consider with all of the studies examined on the basis of
gender is the limited subject pool we have access to for our experiments. All of
our subjects attended Syracuse University, a private university in central New York,
that mainly consists of white students who clearly have access to the privileged
post-secondary educational system. Furthermore, recent trends indicate that the
majority of people who attain advanced degrees in the U.S. are women (Peter &
Horn, 2006). Given that more women sign up for our studies, declare Psychology
as a major, and attend Syracuse University, it is reasonable to expect the women
in our subject pool to be more motivated and focused while completing the exper-
iment. If more women are demonstrating a genuine interest in attaining academic
achievements than men, they may be putting forth a comparatively greater effort
to complete the task. Although we did exclude those subjects who did not properly
complete the task, we were unable to account for individual differences affecting
focus, motivation, and attention.
GENDER & MEMORY 27
Regarding the mathematical distractor task from the delayed free recall ex-
periment, women’s performances may be largely reflective of the stress induced by
stereotype threat. Stereotype threat can alter behavior drastically, provoking per-
sonally harmful behavior associated with the internalization of particularly damag-
ing stereotypes. Even though a woman may have the ability to do well on a math
exam, she may perform below her potential due to the pressures and stress involved
with the actual test, because of the negative constant reinforcement she has received
throughout her entire life, from the media, education system, and the blatant reality
of the workplace, where men overwhelmingly hold a majority of the careers in the
mathematical and scientific domains (Fox, 2001).
To further speculate on the nature of the subtle gender differences we have
observed in cognitive function, we will explore the influence of hormonal fluctuation
in the female brain on memory. Here, we are not assuming that the participants who
identified themselves as “female” in our cognitive studies had e qual levels of estrogen,
or that they necessarily maintained higher levels of estrogen than some subjects who
indicated that they were “male”. However, estrogen is an influential hormone in the
female brain, and the potentially enhancing effects of estrogen on cognitive function
provide reason to consider it as a variable for future research studies. Furthermore,
it is particularly appealing to discover links between behavioral cognitive modeling
and neurological findings in an attempt to determine the underlying causes of various
observations across disciplines.
The hippocampus is the site in the brain where associations among items and
events are formed and episodic memories temporarily stored (Bunsey & Eichenbaum,
1996). Many animal studies have investigated the impact of estrogen on hippocam-
pal structures and processes. In vitro and behavioral studies have unveiled an array
of enhancing effects evoked by the presence of estrogen in the hippocampus un-
der natural and experimental conditions. Unfortunately, the beneficial properties
estrogen apparently induces in the hippocampus of animals have not b ee n directly
GENDER & MEMORY 28
correlated to cognitive studies involving humans. However, due to estrogen’s physio-
logically influential characteristics, it is of great interest to gather implications from
neurological studies involving the impact of estrogen on brain structures believed to
be responsible for basic cognitive functions.
Estrogen has been demonstrated to have an enhancing effect on synaptic plas-
ticity in the hippocampus (Foy, 2001). Synapses are the communicative sites in
the brain where messages are relayed between neurons. Synaptic plasticity refers
to modifications to the formation or structure of the synapse following neuronal
activation. Because of the elongated potency of the synapse following brief synap-
tic activity, researchers believe that synaptic plasticity plays an important role in
memory storage. Cellular changes include an expansion of neural projections and
a subsequent increased number of synaptic connections between neurons. These
resultant alterations are thought to be closely associated with learning and mem-
ory, particularly when neurons within the hippocampus or the cerebral cortex are
affected. Estrogen enhances this process by increasing the number of synaptic con-
nections maintained by a neuron, and thereby potentially improving its ability to
process information (Foy, 2001).
In conjunction with the augmented synaptic plasticity in the hippocampus
provoked by the presence of estrogen, an increase in dendritic spine density on CA1
pyramidal cells is also observed. Dendritic spine density is influenced by the lev-
els of estrogen present during the female rat’s estrous cycle. When estrogen is at
its highest level, during proestrus, spine density is similarly heightened. Concur-
rently, spine density reaches its lowest value during estrus when estrogen levels are
diminished (Woolley & McEwen, 1994). The estrogen-induced increase in spine
and s ynapse density has been positively correlated to working memory performance
in mice (Daniel & Dohanich, 2001), where working memory can be classified in a
synonymous category as short term episodic memory for humans. In a study us-
ing hippocampal slices of ovariectomized rats, following behavioral testing, Daniel
GENDER & MEMORY 29
and Dohanich (2001) observed a parallel increase in NMDA receptor binding and
working memory acquisition in an eight-arm radial maze task following the acute
administration of estrogen 48 or 72 hours before testing.
The excitatory neurotransmitter, glutamate, mediates synaptic transmission
within the hippocampus. Several different receptors bind to glutamate and cause
the neuron to become excited within the hippocampus and provide input to CA1
pyramidal cells, including the NMDA (N-methl D-aspartate) and AMPA (theta-
amino-3-hydroxy-5-methyl-4-isoxazoleproprianate) receptors. Estrogen is presumed
to positively affect the growth of neural projections and enhance synaptic plasticity
through both NMDA and AMPA receptors (Foy et al., 1999). Additionally, other
studies have suggested that NMDA receptor activation is necessary in order to fa-
cilitate the heightened spine density of CA1 pyramidal cells provoked by estrogen
(Woolley & McEwen, 1994).
The magnitude of excitatory postsynaptic potentials (EPSPs) within the hip-
pocampus is intensified by the presence of estrogen. Particularly, AMPA and NMDA
mediated response activity is heightened by estrogen (Foy et al., 1999). An in vitro
study conducted by Foy et al. measured isolated AMPA and NMDA receptor-
mediated EPSPs following the administration of estrogen to CA1 pyramidal cells
located in the hippocampus. Estrogen rapidly enhanced both AMPA and NMDA
receptor channels as evidenced by the increased amplitude of EPSPs evoked by
Schaffer collateral stimulation (Foy et al., 1999).
Long-term p otentiation (LTP) refers to an extended increase in synaptic trans-
mission provoked by brief impulses of electrical stimulation in various neuronal path-
ways (Foy, 2001). Several studies have suggested that the sustenance of LTP may
play an important role in learning and memory (Barnes, 2003). More precisely,
LTP is currently viewed as a potential mechanism of memory storage within the
hippocampus (Foy, 2001)
Within the hippocam pal CA1 region, the induction of LTP requires stimula-
GENDER & MEMORY 30
tion with a frequency that is high enough to sufficiently activate NMDA receptor
pathways. Furthermore, the maintenance and e xpression of LTP is dependent upon
enhanced AMPA receptor function (Foy, 2001). Estrogen has the ability to enhance
synaptic transmission and LTP in CA1 neurons of adult, male rats, as indicated
by the observation of hippocampal slices.(Foy et al., 1999). Additionally, stud-
ies involving awake, female, ovariectomized rats revealed an enhancement of LTP,
either within 20 or up to 60 m inutes, following the administration of estradiol ben-
zoate to the CA1 region. Other in vivo studies have demonstrated the facilitation
of LT P induction to be maximal in female rats during the afternoon of proestrus
(Cordoba Montoya & Carrer, 1997).
The production of granule cells within the adult rat’s dentate gyrus is notably
boosted when estrogen is present. In addition to the granule cell proliferation in-
duced by estrogen, the survival rate of these adult-generated cells is also enhanced.
These findings have been implicated by studies involving the rat’s natural fluctua-
tion of ovarian hormones during their estrous cycle and in studies of animals that
have been ovariectomized and injected with estrogen (Tanapat, Has tings, Reeves, &
Gould, 1999).
Interestingly, female rats actually produce more new granule cells than males.
This sex difference seems to be directly related to the heightened cell proliferation
in the dentate gyrus of the female’s brain during proestrus or immediately following
the administration of estradiol. Production of new cells peaks during proestrus, al-
though they rapidly deteriorate when estrogen levels are naturally minimized during
estrus and correspondingly in cases where the rat has been ovariectomized. Sub-
sequently, even though females generate more granule cells than males, the overall
number of granule ce lls in the dentate gyrus is not significantly differe nt across the
estrous cycle. Specifically, a sex difference in the number of newly generated cells no
longer exists between male rats and female ovariectomized rats that have received
estrogen treatment 14 days prior to comparative testing. Moreover, a greater num-
GENDER & MEMORY 31
ber of newly formed cells degenerate in the dentate gyrus of female rats than males
(Tanapat et al., 1999).
The rapid formation of synapses among newly generated granule cells within
the hippocampus has been implicated to play an important role in memory storage
and retrieval. However, the impact of adult neurogenesis on memory and learning re-
mains controversial. Studies have demonstrated that the survival of adult-generated
granule neurons is enhanced in rats exposed to learning tasks that are dependent
upon the hippocampus (Tanapat et al., 1999). Although the functional significance
of adult neurogenesis is currently unknown, it seems plausible that it is essential to
hippocampal integrity. Since the it is the site in the brain where memories are stored
and proces se d temporarily until they are transferred to other areas of the brain, the
ongoing reproduction of new cells may be a likely contributor to the maintenance
of these hippocampal functions (Tanapat et al., 1999).
Since the level of estrogen in the brain is essential to determine its influence
on the neurophysiology of the hippocampus and its subsequent effect on cognition,
cognitive studies measuring sex differences should assess hormone levels of partic-
ipants at time of testing. The analysis we have provided do e s not account for the
natural fluctuation of estrogen throughout the female menstrual cycle, since pre-
sumably the woman participants were tested at varying times of their cycle, and
other factors that influence estrogen levels (i.e. birth control) were not considered.
However, we can reasonably assume that the woman participants did have overall
higher levels of estrogen than the male subjects. Regardless of the circumstances
in our present study, estrogen levels and resultant hippocampal alterations provide
prospective implications to the study of memory and future treatments available for
cognitive deficits.
REFERENCES 32
References
Barnes, C. A. (2003). Long-term potentiation and the ageing brain. Philosophical Trans-
actions of the Royal Society B, 358 (1432), 765-72.
Bunsey, M., & Eichenbaum, H. B. (1996). Conservation of hippocampal memory function
in rats and humans. Nature, 379 (6562), 255-257.
Cordoba Montoya, D. A., & Carrer, H. F. (1997). Estrogen facilitates induction of long
term potentiation in the hippocampus of awake rats. Brain Research, 778 (2), 430-8.
Daniel, J. M., & Dohanich, G. P. (2001). Acetylcholine mediates the estrogen-induced
increase in nmda receptor binding in ca1 of the hippocampus and the associated im-
provement in working memory. The Journal of Neuroscience, 21 (17), 6949-56.
Fox, M. F. (2001). Women, science, and academia: graduate education and careers. Gender
& Society, 15 (5), 654-666.
Foy, M. R. (2001). 17beta-estradiol: effe ct on ca1 hippocampal synaptic plasticity. Neuro-
biology of Learning and Memory, 76 (3), 239-52.
Foy, M. R., Xu, J., Xie, X., Brinton, R. D., Thompson, R. F., & Berger, T. W. (1999).
17beta-estradiol enhances NMDA receptor-mediated epsps and long-term potentiation.
The American Physiological Society, 81 (2), 925-9.
Friendly, M., Franklin, P. E., Hoffman, D., & Rubin, D. C. (1982). The Toronto Word
Pool: Norms for imagery, concreteness, orthographic variables, and grammatical usage
for 1,080 words. Behavior Research Methods and Instrumentation, 14, 375-399.
Herlitz, A., Nilsson, L. G., & Backman, L. (1997). Gender differences in episodic memory.
Memory & Cognition, 25 (6), 801-11.
Herlitz, A., & Yonker, J. E. (2002). Sex differences in episodic memory: the influence of
intelligence. Journal of Clinical and Experimental Neuropsychology, 24 (1), 107-14.
Howard, M. W., Bessette-Symons, B., Zhang, Y., & Hoyer, W. J. (2006). Aging selectively
impairs recollection in recognition memory for pictures: Evidence from modeling and
ROC curves. (in press)
REFERENCES 33
Howard, M. W., Jing, B., & Provyn, J. P. (Submitted). Bridging the gap: Transitive
associations are formed from learning items in similar temporal contexts. Manuscript
submitted for publication.
Howard, M. W., & Kahana, M. J. (1999). Contextual variability and serial pos ition effects
in free recall. Journal of Experimental Psychology: Learning, Memory, and Cognition,
25 (4), 923-41.
Howard, M. W., & Kahana, M. J. (2002). A distributed representation of temporal context.
Journal of Mathematical Psychology, 46 (3), 269-299.
Johns, M., Schmader, T., & Martens, A. (2005). Knowing is half the battle: teaching
stereotype threat as a means of improving women’s math performance. Psychological
Science, 16 (3), 175-9.
Kramer, J. H., Yaffe, K., Lengenfelder, J., & Delis, D. C. (2003). Age and gender interac-
tions on verbal memory performance. Journal of the International Neuropsychological
Society, 9 (1), 97-102.
Lewin, C., & Herlitz, A. (2002). Sex differences in face recognition–women’s faces make the
difference. Brain and Cognition, 50 (1), 121-8.
Peter, K., & Horn, L. (2006). Gender differences in participation and completion of un-
dergraduate education and how they have changed over time. Education Statistics
Quarterly, 7 (1 & 2).
Primoff, E. (1938). Backward and forward associations as an organizing act in serial and in
paired-associate learning. Journal of Psychology, 5, 375-395.
Pronin, E., Steele, C. M., & Ross, L. (2004). Identity bifurcation in response to stereotype
threat: women and mathematics. Journal of Experimental Social Psychology, 40, 152-
168.
Schwartz, G., Howard, M. W., Jing, B., & Kahana, M. J. (2005). Shadows of the past:
temporal retrieval effects in recognition memory. Psychological Science, 16 (11), 898-
904.
Tanapat, P., Hastings, N. B., Reeves, A. J., & Gould, E. (1999). Estrogen stimulates
a transient increase in the number of new neurons in the dentate gyrus of the adult
female rat. The Journal of Neuroscience, 19 (14), 5792-801.
REFERENCES 34
Woolley, C. S., & McEwen, B. S. (1994). Estradiol regulates hippocampal dendritic spine
density via an N-methyl-D-aspartate receptor-dependent mechanism. The Journal of
Neuroscience, 14 (12), 7680-7.
Yonelinas, A. P. (2001). Components of episodic memory: the contribution of recollection
and familiarity. Philosophical Transactions of the Royal Society B, 356 (1413), 1363-74.
